Переход к жизни в наземной среде на суше и в воздухе, разнообразие форм и способов передвижения в классе рептилий (включающем подклассы черепах, ящериц, змей, крокодилов), связаны с возникновением новых функций и появлением ряда морфологоанатомических изменений тела, структуры органов, развития нервной системы, а вместе с тем образа жизни и поведения животных этой группы.
Мозг рептилий достигает большего развития, чем мозг амфибий, в особенности в отношении увеличения полушарий, обонятельных и зрительных долей, появления серого вещества мозга, коры больших полушарий (Табл. 2.1; 4).
Появление коры означает освобождение мозга от непосредственной связи с рецепторами, здесь возникают широкие возможности переключения, регуляции, обобщения отдельных рецепторных процессов.
В противоположность тому, что наблюдается у амфибий, при удалении даже боковых частей переднего мозга рептилии теряют способность самостоятельно принимать пищу, избегать раздражителей, ранее обращавших их в бегство, в то время как амфибии при удалении большого мозга (без повреждения зрительных долей) выполняют все главные биологические функции — плавание, ловлю добычи, зарывание в землю и даже спаривание.
Наружные покровы тела рептилий защищены от высыхания роговыми чешуйками, что привело к большой утрате чувствительности кожи к химическим, механическим, тепловым и световым раздражителям.
Взамен этого, бо́льшее развитие получили вкусовые органы рептилий, сконцентрированные в полости рта, — обоняние, слух и особенно зрение. По обонянию рептилии находят пищу. Черепахи легко выбирают предпочитаемую пищу из смеси разных сортов питательных веществ. Змеи по обонянию выискивают живую добычу.
У черепах доказано различение черного и белого цвета, дифференцирование рисунков на картонах, расчерченных черными и белыми вертикальными полосами, от картонов, расчерченных горизонтальными полосами того же цвета. Черепахи дифференцируют картон с узкими белыми полосами (в 2 мм) на черном фоне от такового с широкими полосами (в 3—8 мм). Но те же черепахи не различают картонов с изображением белых форм (креста и звезды) на черном фоне[80].
У рептилий сильнее, чем у амфибий, развит слух: они реагируют на тихие, но биологически значимые для них звуки — шелест листьев, шуршание насекомых, копошащихся в траве. Молодые гремучие змеи реагируют на звук в 86 колебаний в секунду, передаваемый через воздух, и на звук в 344 колебания в секунду, передаваемый через почву.
Ящерицы различают звуки гальтоновского свистка от 4600 до 125 000 колебаний в секунду. Ящерицы, испытанные в звуконепроницаемом пространстве, реагировали на звуки 7400—8200 колебаний в сек.[81].
Сами рептилии издают звуки шипения при обороне. У них имеются и позы «обороны».
В соответствии с приспособлением к жизни на земле, под землей, на деревьях и даже в воде у рептилий чрезвычайно разнообразны способы передвижения. Рептилии могут ходить, бегать, ползать, лазать по деревьям, некоторые плавают (крокодилы, водяные черепахи) и в виде исключения даже перелетают сверху вниз планирующим полетом с дерева на дерево (летающий дракон).
Это осложнение организации и многообразие способов передвижения отражаются и на эволюции поведения рептилий, на расширении ориентировочной деятельности, на увеличении их активности при выполнении основных биологических функций, например, питания.
Жизнь в сухопутной среде, изменение характера пищи, привело к возникновению зубов (иногда ядовитых) и преимущественно к хищному способу питания живой мелкой и крупной добычей (начиная от насекомых, червей, моллюсков до крупных животных: рыб, амфибий, птиц и даже млекопитающих, включая низших обезьян). Все это отразилось на осложнении инстинкта питания.
Разнообразие пищи связано с активной комплексной деятельностью рецепторов рептилий при осуществлении акта питания. Например, при охоте за живой добычей, при ее отыскивании, подстерегании, преследовании, нападении, кусании, при поимке сопротивляющейся жертвы и при ее умерщвлении требуется, чтобы у нападающего животного (например, гигантского удава) осуществлялось координированное действие зрения, обоняния, осязания (языком) в соответствии с двигательной активностью (головы, шеи, туловища, всего тела, челюстей) при схватывании, удушении или кусании жертвы. Акт питания состоит из длинной цепи последовательно и преемственно наступающих рефлекторных актов, несомненно, связанных с осуществлением сложных аналитико- синтетических процессов мозга.
Практический анализ происходит у рептилий при дифференцировании многообразных пищевых раздражителей, обладающих различными свойствами по виду, вкусу и другим признакам.
Практический синтез участвует при ответной реакции животного в процессе объединения им ощущений, идущих от отдельных органов, от показаний тех или иных рецепторов и сочетаний этих показаний со сложной двигательной активностью особи.
Инстинкт самозащиты проявляется у рептилий весьма рельефно.
Ящерица ушастая круглоголовка при встрече с опасным для нее животным раскрывает пасть, шипит, подпрыгивает навстречу врагу, при этом у нее по бокам от головы оттопыриваются широкие, розового цвета складки, которые придают ей весьма необычный «устрашающий» вид.
Австралийская плащеносная ящерица (Chlamidosaurus kingi) в случаях опасности не только расправляет большой зубчатый, окаймляющий ее голову воротник, но прижимает к земле заднюю часть тела, поднимает переднюю, вытягиваясь в вертикальном положении, обнажает зубы и потом, если нападающее животное не отступает, переходит в наступление и жестоко кусается.
Инстинкт сохранения потомства у рептилий в общем выражен в той же степени, как и у амфибий, а их способы ухода за развивающейся молодью более или менее сложны, в зависимости от условий. В одних случаях отложенные яйца лишь помещаются в укрытии под камнями, обогреваемыми солнцем, в других — активность материнского организма увеличивается, и она сама вырывает нору во мхе, листьях, во влажных теплых местах; более сложная форма опеки яиц связана с устройством гнезда, т. е. самка настилает слой растений, на которые откладывает яйца и прикрывает сверху отложенные яйца растениями, что обеспечивает возникновение вследствие гниения растений более высокой температуры, необходимой для развития яиц.
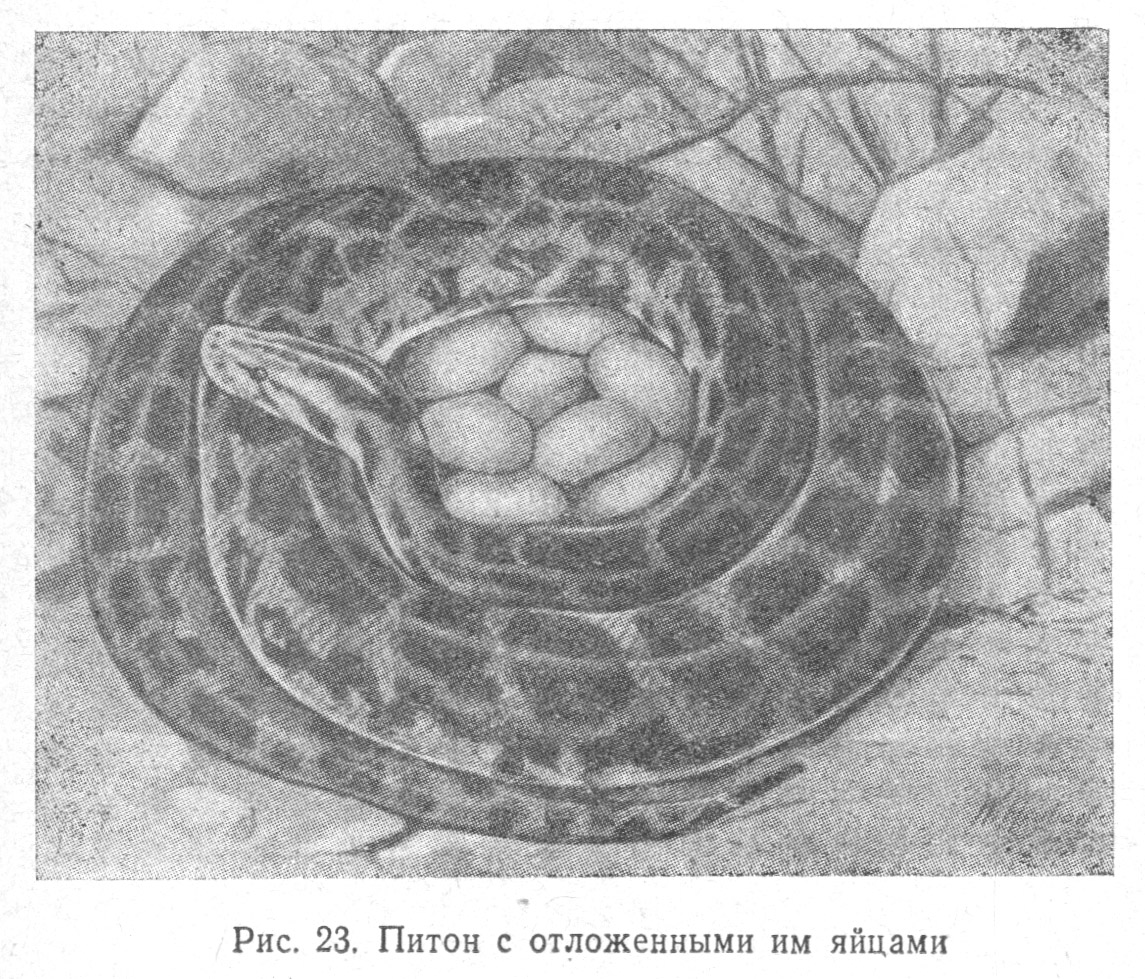
Имеются случаи бо́льшего участия самих родителей в уходе за молодью, обвивание отложенных яиц (Табл. 2.6) своим телом (у питона), насиживание отложенных яиц, охрана гнезда, активная защита гнезда от врагов (бросание на них и кусание) и, что особенно интересно, помощь вылупляющимся детенышам при их освобождении из яйцевых скорлупок. Известны случаи, когда мать сопровождает детенышей при переходе их в ближайший водоем (у крокодилов).
В этих случаях сложная инстинктивная, безусловно-рефлекторная деятельность рептилий предполагает осуществление ряда практических аналитико- синтетических процессов, связанных не только с дифференцированием свойств окружающей среды и активным обеспечением благоприятных условий для созревания яиц, но и установлением контакта с вылупившимися детенышами.
Аналитико-синтетическая деятельность мозга рептилий, связанная с индивидуально выработанными формами поведения, выражена в значительно большей степени, чем у амфибий. Это находит свое подтверждение в том, что содержащиеся в неволе рептилии приучаются различать индифферентные звуковые и зрительные стимулы oт биологически значимых.

Для некоторых ящериц сигнальным стимулом являлся звук, который сначала связывался с едой, а потом и без еды, побуждал их подходить к кормушке с пищей. Для других рептилий вид сосуда, ящика с пищей, появление кормящего человека были сигналом к получению корма, и животные приходили в возбуждение и производили поисковые движения при появлении этих объектов.
Советский физиолог Э. А. Асратян выработал у болотной черепахи условный рефлекс поднятия лапы при подаче звука или вспышке света, предварительно сочетая эти стимулы (свет или звук) с включением тока, проходящего через электроды, прикрепленные к ноге животного. После ряда опытов черепаха стала отдергивать ногу при одной подаче звука или света до момента включения тока. Черепаха могла даже дифференцировать более низкие и более высокие тона, если, например, низкий сочетался с подачей тока, а высокий не сочетался. Прекращение подачи тока приводило к угасанию действия раздражителя, и черепаха совершенно переставала отдергивать лапу (Табл. 2.7)[82].
У ящериц наблюдалась генерализация звуковых раздражителей, и при выработке у них за 7 дней тренировки условного сигнала на тон С (до) они реагировали сходным образом и на тон А (ля).

Установлена ассоциативная память рептилий на места их стоянок, а в условиях эксперимента — способность к запоминанию кратчайшего пути прохождения лабиринта. Водяные черепахи (Clemis gutata) после ряда опытов научились безошибочно проходить кратчайшим путем лабиринт с четырьмя тупиками и выходным свободным концом, который вел к гнезду, затрачивая на прохождение в последнем опыте по сравнению с первым в 60 раз меньшее время, т. е. вместо 35 минут — 35 секунд (Табл. 2.8). В более сложном лабиринте, имеющем шесть тупиков, у черепахи через 50 дней опытов наблюдалось сокращение времени прохождения лабиринта примерно в три раза[83].
Таким образом, у рептилий, по сравнению с амфибиями, ассоциативные связи, относящиеся к синтезу осязательно-кинестетических восприятий, оказываются более совершенными. Сравнение уровня отражения окружающего у рептилий и амфибий указывает, что этот уровень у рептилий значительно повысился. Это повышение связано с увеличением остроты их зрительной и слуховой рецепции и активности по отношению к живым объектам окружающей среды. Напомним активные позы угрозы, связанные с инстинктом самосохранения, активность рептилий в актах питания, в борьбе с живой большой сопротивляющейся жертвой. Несомненно, большая активность рептилий наблюдается и в уходе за молодью в отношении не только вылупляющихся, но и растущих детенышей.
Все эти сложные инстинктивные, как и приобретенные взаимоотношения рептилий, конечно, предполагают наличие тонкого практического анализа и синтеза действующих раздражителей, соответственно с этим уточнение и усложнение аналитико-синтетической деятельности мозга животных, имеющего уже зачаточную кору.
Наличие коры объясняет нам и значительное развитие условнорефлекторной деятельности рептилий, их способность к более адекватным реакциям на зрительные и звуковые сигнальные раздражители.
[80] С. J. Warden, Т. N. Jenkins, L. Н. Warner. Introduction to comparative Psychology. Ronald Press Company. New-York, 1934, p. 464.
[81] F. Hempelmann. Tierpsychologie. Akad. Verlagsgesetlschaft, Lpz., 1926, p. 389.
[82] См. Ю. П. Фролов. Высшая нервная деятельность (поведение) животных. Учпедгиз, 1953, стр. 73.
[83] R. М. Yerkes. Space perception of tortoises, J. Comparat, neurol. and psychol. v. 14, 1904.