Ставится механизм нового типа — горизонтальная задвижка, О (Рис. 1.12), с сильно выступающим в виде диска на столбике мало податливым центром приложения силы, требующим большого активного усилия при отодвигании его в сторону (слева направо) в целях отмыкания (Фото 1.7, Фото 1.8, фиг. 2).
Этот центр приложения силы усматривается обезьяной сразу, но в силу его малой подвижности обезьяна не обнаруживает скоро направления его податливости, а нащупывает его различными способами и в разных местах: она совершает многообразные движения, испытывает всевозможные приемы, концентрируя сначала свое активное внимание по преимуществу на центральной части механизма.
Она захватывает диск в зубы, в рот, тащит вверх и вниз, к себе и в сторону, но в сторону, противоположную направлению отмыкания (именно в левую, а не в правую).
Не преуспевая здесь после 8 минут бесплодной работы, обезьяна обращается, как обычно, к другим частям клетки: то она с силой толкает дверь руками, упирается в нее ногами, налегает на нее всем телом, сотрясает ее, просовывает пальцы в петли сетки, грызет дверь у краев, подсматривает в дверную щель, стучит в стенки руками. Не преуспевая в отмыкании, она обращается опять к центральной части механизма, но все не может сдвинуть задвижку.

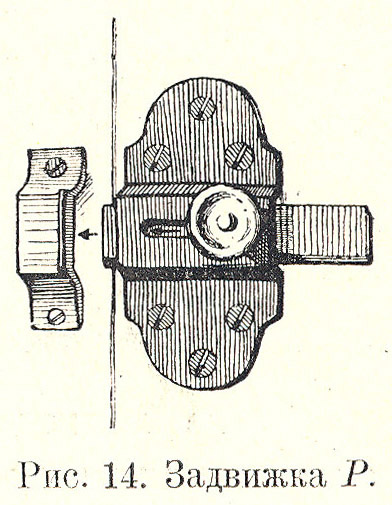
После 23 минут бесплодной работы обезьяна бросает механизм и не желает его трогать; тогда ей предлагают задвижку идентичной конструкции P, лишь более легко податливую в отношении отодвигания центра приложения силы (Рис. 1.13).
Первые опыты и с этой задвижкой безрезультатны. Обезьяна тянет за диск к себе, толкает дверь, налегает всем телом на дверь, сотрясает дверь, просовывает пальцы в петли сетки, грызет углы клетки, подсматривает в щели, грызет близ щели, зубами касается задвижки, опять тянет к себе и, не находя подвижности, бросает работу. Нужное движение не улавливается скоро, хотя обезьяна на этот раз пускает в дело даже зубы; но, ухватив ими за диск, она тащит механизм чаще всего к себе, а не вправо, и конечно наталкивается на совершенную неподвижность механизма; как и всегда, обескураженная неудачей, она после часа работы вообще перестает считать механизм центром задержки, повидимому теряет надежду выбраться из клетки, становится инертной и совсем не желает работать.
Вновь побуждающим к работе стимулом является наглядное выявление связи между отодвиганием задвижки и открыванием дверцы, примерное троекратное отмыкание механизма самим экспериментатором в присутствии обезьяны; открывание дверцы экспериментальной клетки.
Этот маневр сразу воодушевляет обезьяну, и она опять начинает энергично работать над отмыканием.
Вдруг (в 5 сек.) она случайно отмыкает задвижку, но не замечает отомкнутости, так как толкает дверь не сразу, а лишь несколько спустя, после бесцельных манипуляций около того же механизма.
За случайность, как обычно, говорит отсутствие непосредственного толкания двери вслед за отмыканием, наличность нецелесообразных повторных проб в последующем, значительное увеличение времени работы во втором опыте (до 25 сек.), усложненность приема отмыкания. Ухватываясь то зубами, то руками, то опять зубами за выступающий диск, обезьяна отводит центр приложения силы в нужном направлении, двигая голову в правую сторону. Уже в 9-м опыте обезьяна опускает движение руками, продолжая захватывать зубами и передвигать задвижку посредством поворота головы вбок.
Впрочем, и в отношении этого механизма наблюдается с каждым новым опытом резкое улучшение работы, снижение длительности времени отмыкания, рельефно отражающееся на крутом спадании кривой (см. Кривая 1.8). Уже в 10-м опыте мы имеем минимальный срок отмыкания (2 сек.). Среднее время завершения работы в 1-й день — высоко (337,1 сек., т. е. 5,6 мин.).
На 2-й день предъявления того же механизма, после ночного перерыва в работе, наблюдается в 1-м опыте слабое повышение кривой (время отмыкания равно 10 сек.), впрочем быстро понижающейся в последующих опытах. Среди особенностей в манипуляциях обезьяны с этим механизмом следует отметить наличность активного участия зубов в процессе отмыкания (участия, сохраняющегося вплоть до последнего опыта) и полное выключение движений рук.
Эти последние употребляются лишь для контролирования двери, в которую обезьяна упирается одновременно с тем, как производит челюстями отодвигание задвижки в сторону.
Среднее время завершения работы во 2-м сеансе равно всего 5 сек., оно резко укорачивается против такового предыдущего сеанса. Пределы вариации длительности единичных опытов заключаются в периоде времени 3—10 сек. (см. Кривая 1.8). Общая средняя длительность опыта, при учете ее в 2 сеансах, равна 233 сек. (3,8 мин.)
Только после удачного оперирования с задвижкой P предъявляется опять 1-й механизм аналогичной конструкции с более трудно податливым центром приложения силы — задвижка O (см. Фото 1.7, фиг. 2).
Вопреки идентичности приема отмыкания обоих механизмов, обезьяна и теперь, даже познав на опыте прием отмыкания, даже применив нужный прием, ухватив центр приложения силы зубами или руками, — не прилагает достаточного усилия при отодвигании и не преуспевает в разрешении задачи.
Тщетность испытанных проб сбивает животное, отклоняя его внимание с раз испытанного правильного пути — и в результате бесплодных попыток отмыкания обезьяна игнорирует уже и центральное препятствие (механизм), и центр приложения силы в нем (диск на ножке), ищет выхода в других частях клетки, пробует иные приемы работы: то она пытается открыть съемный потолок клетки, то сотрясает, боковые дверцы, то грызет сетку, затягивающую дверцы, то, наконец, она теряет терпение и совсем перестает работать.
В целях стимулирования обезьяны — опять в ее присутствии и при созерцании ею действия — несколько раз производится отмыкание задвижки и открывание двери самим экспериментатором; обезьяна всякий раз после этого тотчас же обращается к работе, но — увы! — еще на меньший срок и еще с меньшим успехом.
То, взявшись верным приемом за отмыкание, она не прилагает достаточно силы при отодвигании, то, не чувствуя податливости в центре приложения силы, она опять начинает его игнорировать, берется за механизм как придется и где попало; с каждым обращением обезьяна манипулирует все более и более кратковременно, все менее охотно; она оперирует с механизмом 1—2 сек., делая длительные паузы между обращениями, уходит в глубь клетки и занимается там «своими делами» — перебиранием шерсти, присматриванием к происходящему вне клетки, протыканием пальцев в сетку клетки.
В целях облегчения задачи производится неполное замыкание задвижки.
После периода работы в 60 сек. обезьяна, наконец, самостоятельно отмыкает механизм, действуя и руками и зубами, но она не учитывает сразу его отомкнутости, а освобождается значительно позднее после случайного нажимания рукой на дверь.
Все же этот 1-й удачный опыт (фактически 7-й пробный опыт)
[22]
является сильно стимулирующим ее к работе, и в последующем (опять при полузамкнутости механизма) она начинает работу с громадной энергией; употребляя прежний прием отмыкания, она все более успешно оканчивает работу, после 20 сек. — во втором
[23]
и в 10 сек. — в 3-м опыте
[24],
причем в последнем случае производится полное замыкание механизма; обезьяна работает, не теряя времени на бесплодные отвлечения в сторону, настойчиво, не отрываясь, держа главный центр приложения силы то руками, то зубами, явственно с усилием добивается его отодвигания вопреки полной замкнутости механизма. Таким образом первое самостоятельное отмыкание вполне замкнутого механизма O осуществляется после 1500 сек. работы (см. Фото 1.8, фиг. 2).
Фототаблица 1.4. Операции с выдвижной дверью
Фиг. 1. Выдвигание двери снаружи
Завершение работы ускоряется после того, как обезьяна опускает бесплодные движения руками (которые, повидимому, слишком слабы, чтобы произвести отодвигание), оставляет лишь прием отодвигания при посредстве зубов, производит отмыкание в три приема
[25],
привстает на ноги для открывания
[26],
контролирует дверь, упираясь в нее левой рукой и открывая мгновенно по отодвигании задвижки. За выключением 1-го, особенно длительного опыта, в последующем время отмыкания не поднимается выше 70 сек.; уже в 5-м опыте работа кончается в 5 сек., но в последующем этот срок не остается постоянным и даже в последнем — 12-м — опыте значительно увеличивается до 25 сек. (см. Кривая 1.8).
Эпизодические повышения длительности работы (до 60 сек.) объясняются в начале сеанса опущением движения, производимого при посредстве челюстей и зубов, и оставлением более слабых движений рук; в конце же сеанса имеет место явное уставание обезьяны, ввиду неподатливости механизма, так как никаких посторонних действий и движений обезьяна не допускает; она работает энергично, усердно, целеустремленно, не отвлекаясь ни на одну секунду, не отрываясь, длительно манипулирует с механизмом вплоть до момента полного его отмыкания. Среднее время окончания работы в 1-м сеансе равно 147,9 сек.
Проверка работы с тем же самым механизмом после 2-дневного перерыва подтверждает только что сказанное.
Обезьяна хорошо помнит прием, сразу, легко справляется с задачей, давая в 1-м опыте отмыкания более краткий срок (3 сек.), нежели в последнем опыте предыдущего сеанса; в течение 10 последующих опытов срок отмыкания варьирует в пределах 3—20 сек. Минимальный срок (3 сек.) повторяется часто, среднее время окончания работы во 2-м сеансе равно 6,5 сек. Общая средняя длительность единичного опыта при учете работы в 2 сеансах равна 80,3 сек.

Фиг. 2. Выдвигание двери изнутри
Фиг. 3. Вылезание обезьяны наружу

Ставится механизм аналогичной конструкции — задвижка Q, по сравнению с предыдущими резко несходный по виду: золотистая, узкая с брусковидным стержнем задвижка с явственно выдающимся (в виде выступающей пуговки на короткой ножке) центром приложения силы; механизм расположен в верху рамки двери, требует движения отодвигания снизу вверх. Механизм легко податлив при отмыкании, но вследствие своего вертикального положения обладает свойством самозамыкаться (Рис. 1.14).
Обезьяна сразу (вопреки удаленности механизма от центра дверцы) замечает механизм и обращается к его отмыканию; она хватается правильно за выступающий центр приложения силы, употребляет привычное движение отодвигания вправо, осуществляя его то руками, то зубами. Так как эти действия не приводят к цели, обезьяна быстро бросает работу с механизмом, обращается к другим боковым дверям клетки, толкает их, стремится выйти. Не получая освобождения, она снова обращается к задвижке, берет ее в зубы, явственно нащупывает по разным направлениям место наименьшего сопротивления. Уловив податливый пункт и направление движения, она перехватывает центр приложения силы руками и производит правильное движение отодвигания, вслед за чем толкает дверь, потратив на 1-е отмыкание 70 сек.
Следующий опыт мало удачен, так как механизм самозамыкается по отмыкании и обезьяна не скоро догадывается координировать действия обеих рук, но ввиду легкой податливости механизма обезьяна повторно энергично движет центр приложения силы вверх и вниз, вверх и вниз, присоединив однажды и движения нажимания на дверь, что и дает ей немедленное освобождение.
Большая пластичность механизма быстро обеспечивает совершенство манипуляции с ним в последующем.
Уже в 7-м опыте обезьяна бросает движения зубами и оставляет лишь движение отодвигания одной рукой вверх, употребляя вторую руку для нажимания при открывании двери.
Координация движений обеих рук становится настолько совершенной, что обезьяна уже в 6—7-м опыте получает почти немедленное освобождение из клетки (длительность работы равна 1 сек.). Пределы колебания срока отмыкания в единичных опытах 1—185 сек. Средняя продолжительность опыта 37,8 сек. (см. Кривая 1.8).

Ставится механизм той же конструкции (Фото 1.7, Фото 1.8, фиг. 2; Рис. 1.15) — задвижка R, но с мало выступающим, эксцентрично расположенным, средне податливым центром приложения силы. Механизм приделывается в горизонтальном положении и вследствие этого требует движения отмыкания, аналогичного таковому, которое уже употреблялось обезьяной при оперировании с задвижками О и Р (т. е. движения отодвигания вправо [27].
На этот раз обезьяна не сразу находит главный центр приложения силы; сначала она производит пробы зубами и руками в иных частях механизма, в середине его. Первое удачное отмыкающее движение осуществляется лишь после длительных манипуляций дерганья центра приложения силы вверх, вниз, влево, после ряда проб, нащупывающих направление отодвигания, ищущих признаков податливости механизма в каком-либо направлении; обезьяна, то, ухватившись за центр приложения силы механизма, сотрясает его, то отводит слегка вправо, то опять задвигает влево; она слышит щелкающий звук замыкающего стержня и начинает действовать более энергично. Едва обезьяна нападает на податливый пункт, она быстро подхватывает направление подвижности и начинает горячо работать в этом направлении вплоть до того момента, когда центр приложения силы доведен «до отказа» и механизм отперт.
Первое удачное отмыкание осуществляется после 240 сек. работы, вслед за чем наблюдается резкое систематическое сокращение времени работы
[28];
уже в 7-м опыте фиксируется прием работы: правой рукой обезьяна отодвигает задвижку, левой — открывает дверь; хотя наравне с этим усовершенствованием впоследствии иногда внедряются бесцельные посторонние обращения обезьяны к второстепенным частям механизма; впрочем, срок окончания работы весьма краток, и уже с 6-го опыта он не поднимается выше 6 сек. Пределы колебания длительности работы от 2 до 240 сек. (см. Кривая 1.8). Среднее время завершения работы равно 29,6 сек.
Ставятся одна вслед за другой задвижки, врезанные внутрь дерева дверцы со скрытым замыканием, с различно податливым центром приложения силы, допускающим то меньшую, то большую легкость передвижения (Фото 1.6, фиг. 4).

Задвижка F черная, погруженная в дерево, центр приложения силы — едва выступающий наружу штифт, легко отодвигаемый влево (Рис. 1.16).
Наблюдается мгновенное постижение механизма: обезьяна в 4 сек. в первом опыте и в 3 сек. во втором опыте производит отмыкание. В последующем вариация срока работы заключена в пределах от 2 до 10 сек.; среднее время длительности опыта 4,4 сек. (см. Кривая 1.8).
Прилаживается задвижка G, почти той же конструкции и внешнего вида, что и предыдущая, но требующая прямо противоположного движения отодвигания (вправо); по легкости отмыкания эта задвижка менее податлива, чем предыдущая, но зато центр приложения силы — штифт — выступает больше (Рис. 1.17).
Механизм длительно не преодолевается обезьяной по двум причинам:
вследствие произведения работы по инерции, т. е. отодвигания центра приложения силы в том же направлении, что и у предыдущего механизма, т. е. влево; вследствие отсутствия достаточного усилия при отодвигании.
Обезьяна в течение 180 сек. (3 мин.) настойчиво оттягивает этот центр приложения силы механизма влево, ухватывается за выступающую точку опоры то руками, то зубами, и хотя и не продвигается ни на иоту в передвижении, все же продолжает тянуть туда же, в левую сторону, не пытаясь изменить направление движения.
Четыре длительных приема отмыкания задвижки, отнявшие 12,6 мин. времени, безрезультатны.
Обезьяна не подхватывает даже данного ей подсказа отмыкания.
Когда я полузавожу задвижку и создаю ей условия движения в двух направлениях (вправо и влево), обезьяна останавливается упорно на движении влево, хотя оно снова многократно приводит ее к тупику, ибо этим движением она не отмыкает, а замыкает механизм. Тем не менее она предпочитает даже бросить работу и сидеть, скучая и томясь взаперти клетки, нежели самостоятельно испытать новое направление передвижения центра приложения силы.
Сила инерции столь же препятствует удачной работе, как и некоторая затрудненность отодвигания.

И это доказывается явственно при предъявлении нового механизма Н — задвижки почти аналогичной конструкции, требующей того же направления движения, что и задвижка G (т. е. вправо), лишь с несколько более выступающим (округлым), с более податливым, хотя и слегка пружинящим центром приложения силы (Рис. 1.18).

И в данном случае обезьяна в течение 270 сек. непрерывно оттягивает центр приложения силы в ту же левую сторону, движет то пальцами рук, то зубами.
Случайные обстоятельства, как и всегда, помогают обезьяне: устав оперировать левой рукой, она меняет руку и начинает действовать правой; тогда она невольно нападает на направление податливости механизма в правую сторону, толкает центр приложения силы вправо, задвижка слегка подается и щелкает; обезьяна начинает манипулировать энергичнее, шатает задвижку взад и вперед, вправо и влево то одной, то другой рукой, потом, отодвигая одной правой рукой центр приложения силы, она быстро подхватывает направление передвижения и отводит центр приложения силы так далеко, что достигает отмыкания; одновременное движение толкания двери левой рукой дает ей немедленное освобождение из клетки после 270 сек. работы.
Удачный прием работы запоминается с одного раза, правда, в следующем опыте обезьяна еще сохраняет способ схватывания обеими руками за выступающий центр приложения силы, но скоро, впрочем, она оставляет для манипуляций одну правую руку и получает освобождение в 38 раз скорее, чем в предыдущем опыте.
В последующем обезьяна уже в 4-м опыте сразу начинает оперировать одной правой рукой, ловко отодвигая выступающую пуговицу — центр приложения силы — тремя первыми пальцами, нажимая на дверь левой рукой во все время отмыкания и в 3 сек. кончая работу. Длительность завершения работы в 5-м минимально коротком опыте равна всего 2 сек.; по сравнению с первым опытом уменьшение времени в 135 раз; среднее время завершения работы равно 57,8 сек. (см. Кривая 1.8).
Кривая 1.8. Задвижки P, O, Q, R, F, H, I, G

Кривая 16. Задвижка P, горизонтальная. |
Кривая 20. Задвижка F, штифтовая. |
Предлагается задвижка совершенно иная по виду и по степени заметности центра приложения силы, но тождественная с предыдущей по способу и направлению отмыкания, именно — задвижка I, погруженная в желобок с миниатюрным, открытым, легко отодвигаемым вправо, центром приложения силы, имеющим вид штифта (Фото 1.7, фиг. 1, 2; XI, фиг. 2; Рис. 1.19).

Обезьяна сразу правильно находит центр приложения силы, вопреки его малой заметности и, действуя по инерции с предыдущим, быстро производит нужное движение уже в 1-м опыте в кратчайший срок (2 сек.), получая освобождение и сохраняя совершенство отмыкания этого механизма на все последующее время. Пределы колебания длительности отмыкания — от 1,5 до 3 сек. Средняя длительность опыта 2,2 сек. (см. Кривая 1.8).
Теперь снова ставится задвижка G, имеющая трудно податливый, но отодвигаемый в ту же правую сторону центр приложения силы.
И на этот раз обезьяна вначале не прилагает достаточного усилия, чтобы отодвинуть центр приложения силы, хотя она и берет правильное направление отодвигания и работает в течение 160 сек. Причина неуспеха кроется не в том, что у обезьяны недостает силы для отмыкания задвижки, а в том, что она не производит длительного отодвигания.
Если обезьяна, уцепившись за центр приложения силы и едва потянув его, сразу не ощущает продвижения задвижки — она в ту же секунду отрывается от работы, ища других путей выхода, после чего опять обращается к механизму па столь же короткое время.
Только после 1131 сек. (18,8 мин.), после 9 пробных опытов отпирания, беря центр приложения силы то руками, то зубами, обезьяна производит 1-е самостоятельное отмыкание механизма; она быстро прогрессирует в работе, и уже в 11-м опыте усваивает совершенно точный прием отмыкания, а в 6-м опыте дает минимальный срок отмыкания — 1 сек.
Среднее время завершения работы — 198,6 сек. (см. Кривая 1.8).

Новым, предъявленным для отмыкания механизмом являлась задвижкa Y со скрытым, замаскированным, опущенным в глубокий канал, эксцентрично расположенным чашевидным центром приложения силы, требующим от обезьяны наблюдательности при его отыскании, тонкого движения пальцев при его взятии и известного усилия при его отодвигании. Направление отодвигания было иное, чем у предыдущих механизмов H и I, т. е. на этот раз влево, а не вправо (Фото 1.8, фиг. 1; Рис. 1.20).
В целях отыскания податливого пункта обезьяной применяются многообразные ухищрения.
Обезьяна трогает сверху, снизу, с боков выступающую на дверце задвижку; она кусает, сотрясает дверь, дергает ее к себе, отходит; после 50 сек. работы опять тянет задвижку вправо за выдающуюся часть, но недостаточно энергично; не находя скоро пункта податливости, макак уходит, но, не видя других способов выхода из клетки, опять возвращается; обращается к более тонкому обследованию механизма: тянет за внутренние края чашевидного центра приложения силы, пощипывает за него пальцами, но мало энергично, пристально приглядывается сверху, подглядывает снизу под механизм, но дольше и больше, и прежде всего оперирует при посредстве осязательных проб пальцами, губами, зубами — грызет, кусает, дергает, потом тянет пальцами близ, около, поверх задвижки; потом отвлекается, обращается кверху, толкает потолок, опять кусает задвижку; вдруг одним из движений пальцев, засунутых внутрь задвижки, обезьяна отодвигает до конца за край центра приложения силы и отмыкает механизм через 165 сек. от начала опыта.
Этот первый случайно удачный опыт не научает животное отмыканию, ибо в последующем остаются те же многочисленные нащупывающие движения, лишь более сконцентрированные во внутренней части механизма.
Обезьяна запоминает место задержки, но не улавливает подвижного пункта центра приложения силы, она не усвоила способа и направления его отведения: вместо двигания вправо за край чашевидного центра приложения силы, обезьяна кусает задвижку, просовывает пальцы внутрь, ковыряет, колупает ее близ центра приложения силы, грызет зубами, трогает, тянет за край желоба в левую сторону, кусает центр приложения силы снизу, сотрясает дверь, трогает задвижку то с одного, то с другого конца, дергает слегка за край; или она просто хватает за центр приложения силы пальцами, кусает зубами, или если и дергает его в сторону, то так слабо и кратковременно, что не отодвигает до степени отмыкания.
После 900 сек. (15 минут) бесплодной работы обезьяна начинает все более отвлекаться и бездействовать и, наконец, совсем отказывается работать.
Задача облегчается — задвижка полузамыкается. Этим самым главный центр приложения силы придвигается из эксцентричного в более центральное положение и потому имеет два направления передвижения.
Обезьяна быстрее, чаще обращается к нему в своих операциях; теперь она скоро улавливает его податливость, но совершает движение в прямо противоположную сторону (вправо), т. е. замыкает задвижку; до 15 раз производится полузамыкание задвижки, и столько же раз обезьяна производит то же стереотипное движение замыкания, не ведущее к цели и не дающее ей никаких выгод.
Делается новое облегчение: затрудняется движение центра приложения силы в несоответствующую сторону.
После безрезультатных попыток отодвигания в прежнюю сторону обезьяна утрачивает и последний податливый пункт, теряется окончательно и совсем не желает работать. От времени до времени она делает центром своего внимания как раз противоположный конец задвижки, где видит передвижение выступающего конца стержня.
В виду полного игнорирования обезьяной центра приложения силы приходится несколько раз особенно указывать ей на него, приходится специально поощрять обезьяну за каждое случайное к нему прикосновение.
Теперь обезьяна охотнее обращается к нужной точке, энергичнее, многообразнее манипулирует близ нее, точнее, тоньше берется за работу.
Одним из движений обезьяна вдруг слегка тянет за внутренний край чашки; отодвигая ее в нужную сторону, — тотчас же она получает поощрение; тогда она еще энергичнее работает в той же точке — и быстро достигает удачи, отодвигая до конца за край погруженной в желоб чашки замыкающий стержень.
Обезьяна отмыкает задвижку, потратив в общем на работу 15 минут.
И 2-й удачный опыт
[29]
еще не учит многому; в дальнейшем опять наблюдается и задвигание задвижки в противоположную сторону, и масса нецелесообразных проб, вроде колупанья пальцем, грызенья задвижки за край чашки в желобе; правильное резкое энергичное отодвигание, дающее отмыкание задвижки, производит впечатление случайного действия; оно осуществляется в срок 120 сек.
Только с 4-го опыта обезьяна сразу производит сильное отодвигающее движение, и тогда время завершения работы резко сокращается (до 5 сек.), хотя нецелесообразные движения, вроде грызенья близ свободного конца задвижки, еще вкраплены.
В 5-м опыте задвижка совершенно замыкается.

Обезьяна производит 2-е самостоятельное отмыкание вполне замкнутой задвижки в результате 1249 сек. (20 мин. 49 сек.) предварительной работы; прием работы фиксируется, хотя осложнен, — акт отодвигания осуществляется в 3 приема: после 1-го приема отведения обезьяна контролирует дверь; не получая освобождения, она на этот раз не ищет иных путей выхода, а вторично и третично производит движение отодвигания. Этот прием остается до 11-го опыта.
Только в 14-м опыте вырабатывается прием отмыкания задвижки одним сильным движением отодвигания одним указательным пальцем, что резко сокращает время работы (до 5 сек.); не только в этом опыте, но и во всех последующих длительность отмыкания не более 6 сек. Средняя длительность опыта равна 88,7 сек. (см. Кривая 1.9).
Эпизодическое увеличение времени работы падает не столько за счет оперирования с задвижкой, сколько за счет манипуляций с дверцей, которая, вследствие плотности вхождения в рамку, требует иногда большого усилия для открывания.
Механизм Y со средне податливым, но замаскированным, скрытым центром приложения силы, имеющим направление передвижения обратное таковому предыдущего механизма, не постигается обезьяной самостоятельно, а с подмогой, с „подсказом" экспериментатора. Первый случайно удачный способ отмыкания механизма обезьяной не запоминается. Только после 10 пробных опытов фиксируется определенный способ отмыкания, сохраняющийся на все последующее время.
Теперь обезьяне предъявляется для отмыкания серия механизмов-задвижек, именно 3 задвижки: 1) R (с эксцентрично расположенным, средне податливым, средне заметным центром приложения силы); 2) O (с центрально расположенным, легко заметным, трудно податливым центром приложения силы); 3) I (с центрально расположенным, скрытым, легко податливым центром приложения силы) [30]. (Фото 1.7, Фото 1.8, фиг. 2.) В двух задвижках (R и O) замыкающий стержень открыт, и скольжение его явно заметно, а в третьей задвижке (I) он закрыт. Первоначально замыкается одна верхняя задвижка R, отмыкающаяся движением отодвигания влево. Контролируется способность обезьяны к опознаванию по виду замкнутой задвижки от незамкнутой.
На основании 15 опытов вскрывается совершенно явственно, что первоначально обезьяна не учитывает замкнутости верхней задвижки (R); она манипулирует и с средней (O) и с нижней (I) задвижками, делая большое количество (до 6) излишних движений; но уже в 3-м опыте она перестает обращаться к отмыканию нижней и значительно сокращает излишние прикосновения к средней, касаясь ее не более 1 раза и тотчас же переходя к отмыканию замкнутой верхней задвижки (R). Следует отметить, что это прикосновение к другим запорам, в особенности к средним, осуществляется вследствие того, что обезьяна не прилагает сразу достаточно ловкого и сильного приема при отодвигании верхней задвижки (R); едва коснувшись, потянув за слабо выступающий центр приложения силы, обезьяна уже перескакивает к обследованию 2-го механизма; едва коснется его — и снова возвращается к 1-му, который и отмыкает более настойчиво и окончательно. Едва раздается звук отщелкивания задвижки R — и обезьяна тотчас же толкает дверь, освобождается.
Если обезьяна сразу употребляет правильный прием отмыкания, прилагает достаточно силы и ловкости при отодвигании 1-й задвижки, — она быстро (1—2 сек.) справляется с отмыканием и не обращается к другим запорам; но это имеет место лишь в 25% случаев и не ранее чем в 8 опыте.
Когда в 3 последних контрольных опытах задвижка R не замыкается, обезьяна как будто учитывает эту незамкнутость: она или сразу толкает дверь или, слегка коснувшись задвижки одной рукой, едва взглянув на механизм, уже тянет, открывает дверь второй рукой; но когда после нескольких опытов незамыкания верхней задвижки (R) последняя снова замыкается, обезьяна, отвыкнув от движения отмыкания, опять не учитывает по виду замкнутости; она снова не хочет сразу приложить достаточного усилия и отодвинуть задвижку и потому опять бросается к отмыканию среднего, нижнего механизма, возвращаясь к более настойчивой работе с первым, верхним запором лишь после безуспешной манипуляции с двумя последними механизмами. Среднее время завершения работы (17,8 сек.) несколько меньше такового при первоначальном манипулировании с той же задвижкой, когда она была единичной (там это время было равно 29,6 сек.). Длительность единичных опытов заключается в пределах между 1—81 сек. времени (см. Кривая 1.10).
При замыкании одной нижней задвижки I наблюдается следующая картина работы обезьяны.
Около 5 сек. обезьяна бьется над отмыканием незамкнутой верхней задвижки (R), затем прикасается зубами к отомкнутой средней (O), в течение 30 сек. оперирует с ней, затем пытается отпирать то верхний, то средний механизмы, закрывает и открывает верхний, ухватывается зубами за центр приложения силы среднего, снова настойчиво обращается к верхнему, рвет его, тащит к себе, ломает, принимается снова за средний и привычным маневром (зубами) тянет его к себе; вдруг она обращается, подойдя издали, к нижней задвижке (I), движет ее вправо и влево, но недостаточно энергично; по истечении 3 минут от начала опыта обезьяна еще раз отмыкает и замыкает задвижку, опять отмыкает ее, но забывает контролировать дверь; затем после 5 минут (300 сек.) общего количества работы, наконец, отмыкает и ее и тянет дверь к себе.
При замыкании 2 задвижек — средней O и нижней I (из 3 задвижек R, O, I) наблюдается следующая картина работы.
В течение 5 опытов по инерции с предыдущим обезьяна неизменно обращается к манипулированию с верхней (1-й), на этот раз отомкнутой задвижкой R, с которой она оперирует даже чаще и дольше, чем со всеми другими задвижками; она длительно не усматривает более эксцентрично расположенную 3-ю задвижку (I). Не наблюдается никакого определенного порядка в отмыкании задвижек.
Только 1-й опыт отмыкания обращает на себя внимание, как по сравнительно большому количеству движений, так и по длительности (61 сек.); в последующих экспериментах то и другое резко уменьшается, но, как и обычно при оперировании с комбинацией механизмов, нет систематического спуска кривой времени завершения работы.
Среднее время продолжительности работы по отмыканию 2 механизмов (равное 33,8 сек.) на 49,5 сек. меньше суммы (т. е. 82,3 сек.) среднего времени отмыкания обеих задвижек O и I при предложении их порознь.
Причина этого удлинения — наличность первой задвижки R, по инерции надолго (до самого последнего опыта) приковывающей внимание обезьяны.
У обезьяны наблюдается быстрое образование прочных двигательных навыков, приводящее к автоматизму в действовании, — привыкнув к отмыканию верхней задвижки R, она длительно не обращается к отмыканию нижней I, и средней O, оперируя по инерции с незамкнутой верхней R; отвыкнув от отмыкания задвижки R, обезьяна не сразу обращается к ее отпиранию и после замыкания R; отсутствие зрительного опознавания отомкнутости и замкнутости механизмов является также тормозом в работе.

При предъявлении животному комбинации из двух задвижек О и P, приделанных в том же горизонтальном положении на близком расстоянии (4,5 см) одна от другой (Рис. 1.21), обнаруживаются следующие особенности работы, легко прослеживаемые при рассмотрении главных слагаемых работы в помещенной ниже таблице.
Первый опыт отмыкания весьма кратковременен (10 сек.), но в последующем наблюдается громадная вариация времени работы, в пределах от 4 и до 254 сек.
Среднее время завершения работы с обоими механизмами (38,8 сек.) значительно ниже суммы средней продолжительности работы с каждым механизмом в отдельности
[31],
и это указывает на усовершенствование обезьяны в деле отмыкания 2 механизмов.
Если обезьяна следует определенному порядку при отмыкании, к чему она сама привыкает (т. е. сначала отмыкает нижний механизм P, потом верхний O), то она в среднем не тратит на отмыкание более 14 сек.; если же она утрачивает этот порядок (начинает с отмыкания верхней задвижки), то она часто забывает о нижней задвижке, обращается к другим путям выхода — кверху, к задней дверце, снова к верхней задвижке, отвлекается, иногда вторично замыкает и вторично отмыкает этот верхний механизм и потом уже, после ряда бесплодных попыток выхода, обращается к отмыканию нижнего механизма.
Все эти излишние операции весьма удлиняют время работы (см. 7-й и 9-й опыты).
Эта эпизодическая длительность работы в единичных опытах (равная в максимуме 254 сек.) и делает чрезвычайно широкой амплитуду колебания времени завершения работы (см. Кривая 1.11).
В общем же процент идеальных опытов
[32]
значительно выше среднего, а процент излишних движений весьма не велик. Опытов с отсутствием лишних движений значительно больше, чем опытов с наличностью таковых (см. табл.).
Излишние операции падают главным образом на 2-й механизм именно ввиду того, что, не преуспевая иногда в отмыкании первого, менее податливого механизма (O), обезьяна многократно ищет во втором (P) причину задержки. Обезьяна усваивает определенный порядок отмыкания: как явствует из таблицы, она идет в отмыкании снизу вверх и неизменно сбивается, теряется, удлиняет время работы, если меняет порядок отмыкания — начинает сверху; тогда она зачастую забывает об отмыкании нижнего более удаленного механизма, отвлекается, производит излишние операции, бесцельные движения.
Зрительного различения отомкнутости и замкнутости механизмов нет: обезьяна длительно (секунд 40) продолжает иногда отодвигать задвижку O даже после того, как механизм перестает держать дверь запертой — она двигает центр приложения силы до отказа и иногда вторично замыкает уже отомкнутый механизм P.
Работа задерживается иногда вследствие опускания обезьяной движения контролирования двери, — движения, дающего ей сигнал к высвобождению в случае окончания работы, — к продолжению действий в случае незавершенности отмыкания.

Специальная 20-кратная проверка наличности различения обезьяной отомкнутого механизма от замкнутого дает следующие результаты:
В случае замыкания одного верхнего механизма (O) обезьяна тем не менее, отомкнув его, обращается и к отпиранию отомкнутого нижнего; иногда она даже начинает отмыкание с отпертого нижнего механизма [33].
В случае незамкнутости обоих механизмов обезьяна тем не менее в течение 15 опытов неизменно касается нижнего механизма, прежде чем начинает контролировать отомкнутость двери.
В случае замкнутости одного нижнего механизма обезьяна не касается отомкнутого верхнего лишь потому, что первая же манипуляция с этим механизмом (вследствие одновременного нажимания на дверь) дает обезьяне выход из клетки.

Замыкаются одновременно две другие задвижки (F и G), прикрепленные одна над другой, из которых нижняя (G) отмыкается движением слева направо, а верхняя (F) — движением справа налево. Первый механизм (G), как известно, трудно податлив для отведения, второй (F) более податлив (Рис. 1.22).
Обезьяна в первом же опыте преуспевает в отмыкании обеих задвижек, но она тратит на это отмыкание громадное количество времени — 312 сек., производит 20 обращений к механизмам, прежде чем окончательно справится с задачей. Причина затруднения — это 1-я задвижка (G), требующая большого усилия при отодвигании, — обезьяна не прилагает сразу этого усилия, не получает освобождения и поэтому обращается многократно к повторному оперированию со 2-й задвижкой (F), замыкает и снова отмыкает ее; не получая освобождения, она употребляет несоответствующие приемы отмыкания: дотрагивается зубами, грызет у краев, скребет и, наконец, ухватывается зубами за штифт то той, то другой задвижки, и все же не может отомкнуть; тогда она бросает работу, отходит, снова возвращается и, наконец, достигает освобождения после длительных манипуляций (после 5 мин. 12 сек. работы).
Одного опыта отмыкания достаточно для того, чтобы в последующем побудить обезьяну к более энергичному отмыканию и первой задвижки, но является новое осложнение. Обезьяна после длительных оперирований с механизмами зачастую забывает контролировать дверь и не выходит из клетки, хотя справилась с отмыканием обеих задвижек; не понимая причины задержки и не учитывая отомкнутости, она воспроизводит опять бессмысленные операции грызения этих механизмов, увеличивая без надобности время завершения работы.
Но уже начиная с 4-го опыта она резко сокращает количество излишних движений и соответственно этому и время работы, употребляя правильный прием отмыкания обоих запоров, не забывая своевременно толкать дверь; уже в 8-м опыте она справляется с отмыканием обоих механизмов в минимальный срок (2 сек.).
Все же предшествующие неудачные манипуляции отражаются неблагоприятно на общем итоге опытов (см. табл.).
Среднее время завершения работы очень велико (100,8 сек.), значителен и процент излишних движений (77,1%), велика и амплитуда колебания между максимумом и минимумом времени работы (от 2 до 375 сек.).
Процент идеальных опытов весьма невелик (37,5%), и среднее время завершения работы в этих последних опытах (равное 6 сек.) в 3 раза больше возможного кратчайшего срока завершения работы (2 сек.) (см. Кривая 1.12).
При учете частоты распределения излишних прикосновений приходится сказать, что они почти равномерно распределены между 1-м и 2-м механизмами.
При учете порядка отмыкания механизмов обнаруживается, что обезьяна наичаще последним отмыкает верхний механизм (F), часто она начинает именно с него отмыкание; реже она начинает отмыкание с нижнего (G), но редко и кончает отмыкание этим последним.
Обезьяна идет при отмыкании в направлении сверху вниз и затем опять наверх.

При замыкании двух других задвижек — Y и I, из которых первая расположена вверху дверцы и отмыкается движением справа налево, вторая — внизу дверцы и отмыкается движением слева направо, в результате 30 опытов обнаруживаются следующие особенности работы обезьяны (см. Фото 1.8, фиг. 1, 2; Рис. 1.23; Кривая 1.13).

Излишние движения несколько преобладают над необходимыми (первых — 55,3%, вторых — 44,6%). В особенно неудачных случаях обезьяна производит до 7 ненужных операций, многократно повторно то замыкая, то отмыкая механизмы. Процент идеальных опытов (где эти лишние движения отсутствуют) весьма невелик — именно 16%; следовательно, в 84% опытов (и даже в самых последних) обезьяна производит эти бесцельные операции (см. табл.).
В громадном большинстве случаев эти излишние манипуляции приходятся на долю верхнего механизма (Y)
[34]
и обусловливаются тем, что обезьяна забывает о наличности удаленного из поля зрения нижнего механизма (I) и многократно оперирует с верхним; только после повторного
[35]
контроля двери она обращается к открыванию нижнего запора.
Фототаблица 1.5. Оперирование с крюками
Фиг. 1. Предъявление комбинации из 8 крюков
Излишние операции с нижним механизмом, составляющие только 24,2% общего количества излишних обращений к механизмам, обусловливаются тем, что обезьяна не всегда сразу и удачно отмыкает этот второй механизм и иногда при неумелом взятии за штифт (центр приложения силы) не может отвести его до конца — он застревает на полпути: тогда она бросает работу, обращается к отмыканию другого, верхнего механизма, контролирует дверь, а после того возвращается опять к нижнему, который и отмыкает до конца. Иногда обезьяна при спешной работе не сразу отмыкает до конца и верхний механизм, хотя она в совершенстве владеет приемом его отмыкания.
Верхняя задвижка (Y) не только наичаще привлекает к себе внимание обезьяны, но она и отмыкается (как это видно по таблице) в первую очередь (в 80% случаев она отмыкается первой); она отмыкается первой в 4 раза чаще, чем вторая — нижняя — задвижка. Эта же последняя наичаще отмыкается второй по порядку и еще чаще — последней.
Обращаясь к анализу времени завершения работы с обоими механизмами, следует подчеркнуть, что среднее время завершения работы (равное 15,3 сек.) значительно меньше суммы
[36]
среднего времени завершения работы с теми же, но единично представленными механизмами, и это явно свидетельствует о том, что обезьяна усовершенствовалась в приеме отмыкания обеих задвижек.
Действительно, эпизодическая чрезмерная длительность отмыкания (см. в таблице цифру максимума времени окончания работы), как уже было упомянуто, обусловлена у обезьяны не столько незнанием приема отмыкания, сколько забыванием ею о наличности второго центра задержки — удаленного нижнего механизма.
В удачных случаях завершение работы с обоими механизмами весьма кратковременно (2,5 сек.), но, как это явствует из той же таблицы, среднее время завершения работы даже в идеальных опытах больше этого минимума (на 7,1 сек.), следовательно обезьяна даже в этих так называемых идеальных опытах не работает идеально совершенно — и если она не производит излишних операций, то все же и не кончает работу в тот кратчайший срок (2,5 сек.), в который она могла бы окончить дело. Она тратит на отмыкание в среднем в 3 раза больше времени, чем могла бы; она употребляет в каждом рядовом опыте времени (в среднем) на 5,7 сек. больше того, которое затрачивается (в среднем) на идеальный опыт и которое требовалось бы как необходимое для отмыкания (при условии отсутствия лишних движений).
Насколько велик диапазон вариации времени завершения работы, об этом дает представление сравнение максимума и минимума времени окончания работы. В особенно неудачных опытах работа заканчивается в более долгий (почти в 40 раз) срок, чем в опытах удачных.
В общем и при работе с этими 2 механизмами обнаруживается, что обезьяна дает необычайно низкий процент идеальных опытов, что в среднем даже в идеальных опытах она работает дольше того, чем могла бы; и это происходит главным образом потому, что она забывает об удаленном нижнем механизме.
Обезьяна придерживается порядка отмыкания в направлении сверху вниз.
Фиг. 2. Отпирание крюка e2
Фиг. 3. Отпирание крюка e3

При замыкании двух других задвижек — I горизонтальной, отмыкающейся движением направо, и P1 вертикальной, отмыкающейся новым движением снизу вверх, — результаты работы снижаются по успешности еще более значительно (Фото 1.7, фиг. 2).
Быстро отомкнув горизонтальную задвижку (I), обезьяна контролирует дверь, но, не получая освобождения, она вторично, многократно обращается к замыканию и отмыканию верхней задвижки (I) и не замечает нижней (P1).
Наконец она обращается к задвижке P1, в два приема отмыкает ее, но теперь забывает толкнуть дверь, а потому опять не получает освобождения. Тогда снова обезьяна бесцельно замыкает нижнюю задвижку, после чего бессмысленно контролирует дверь, толкая ее; еще более бесцельно и бессмысленно она на этот раз замыкает и верхний механизм I. Наконец после 330 сек. работы она последовательно отмыкает и 1-й и 2-й механизмы, получая освобождение.
В следующем опыте отмыкания количество излишних движений резко уменьшено — до 8; сокращено время отмыкания до 97 сек.
В 3-м опыте обезьяна почему-то вначале производит вслед за отмыканием нижнего механизма повторное его замыкание, а позднее длительно забывает о нем, а еще позднее, когда вспоминает о нем, употребляет недостаточно энергичный прием отмыкания, и хотя сокращает количество излишних движений до 5, но увеличивает опять время завершения работы до 330 сек.
Среднее время окончания работы чрезвычайно удлиняется (до 253 сек.), по-видимому вследствие того, что механизмы расположены в разных плоскостях, имеют разные направления движения и удаленный механизм P1 длительно не обращает на себя внимания обезьяны; иногда же обезьяна совсем забывает о его наличности, иногда она не сразу улавливает правильный прием его отмыкания, не знает, не учитывает смысла отмыкания, и сама осложняет себе работу повторным замыканием задвижек.
Среднее время завершения работы тем больше, чем эксцентричнее удален из поля зрения обезьяны 2-й механизм и чем новее способ его отмыкания.




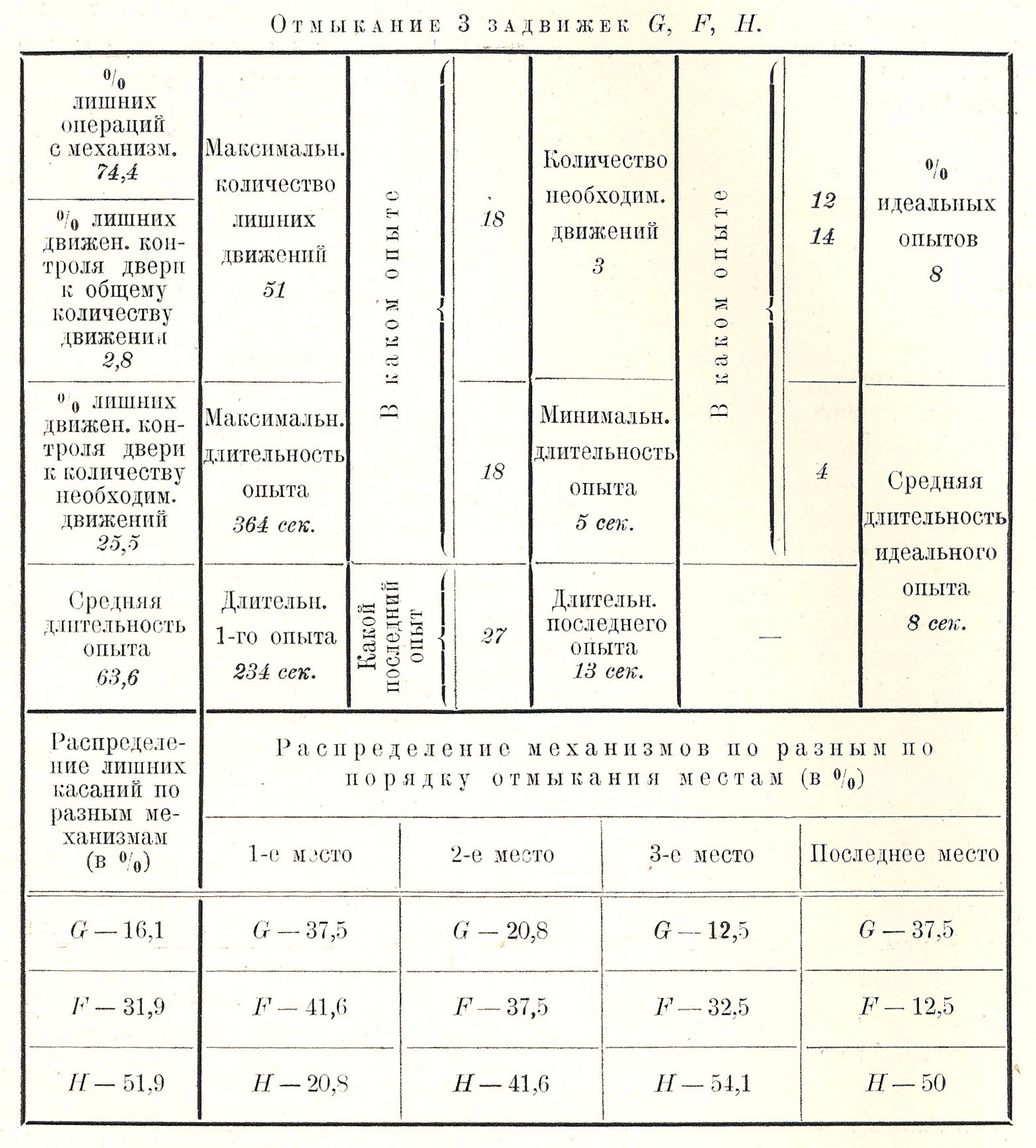
При замыкании 3 штифтовых задвижек (G — нижней, отмыкающейся движением слева направо, F — средней, отмыкающейся движением справа налево, H — верхней пружинящей, отмыкающейся движением слева направо) результаты работы близки к предыдущим (см. табл. и табл. при работе с двумя штифтовыми задвижками G и F и с тремя — G, F, H), хотя и варьируют в деталях (Рис. 1.22).
Во-первых, ввиду предшествующего упражнения в отмыкании 2 задвижек (F и G) среднее время завершения работы значительно (на 40 сек.) меньше, чем при работе с двумя механизмами: оно равно 63,6 сек.; меньше (на 11 сек.) и максимум времени завершения работы в неудачных опытах, но ввиду наличности одного лишнего (3-го) механизма (H) и минимум времени в удачных опытах, и среднее время завершения работы в идеальных опытах несколько увеличены, а количество идеальных опытов значительно уменьшено, почти на 30%.
Количество излишних операций несколько понижено (на 2,4%) по сравнению с предыдущим, но максимальное количество этих операций в единичных опытах возрастает до громадной цифры (51).
Это последнее обязано наличию самозамыкающейся (3-й) задвижки H, которую первоначально обезьяна отмыкает в первую очередь
[37]
и которую она вынуждена зачастую многократно отмыкать ввиду повторного ее замыкания.
Часто обезьяна не учитывает истинной причины задержки (3-й задвижки H) и поэтому по нескольку раз производит обследование двух других задвижек — то отмыкает, то замыкает их; часто обезьяна не производит достаточно энергичного отмыкания задвижек — и, не окончив открывание одной, бросается ко второй, к третьей задвижке и опять к первой; еще чаще обезьяна подряд 3—4 раза в несколько приемов отмыкает одну и ту же задвижку.
Конечно, в силу вышеуказанных причин максимальный процент излишних обращений падает на задвижку H (в 51,9% случаев), во много раз меньше обращение к средней F (в 31,9% случаев) и особенно к нижней задвижке G (в 16,1 % случаев).
По порядку отмыкания первое место занимает наичаще центрально расположенная средняя задвижка F, реже — G и еще резке — верхняя H. Верхняя задвижка H чаще других отмыкается 2-й по счету, тогда как она должна отмыкаться всегда последней, но она отмыкается последней главным образом в конце сеанса и лишь в 50% опытов; нижняя задвижка G равно часто отмыкается и первой и последней.
Таким образом чаще всего обезьяна начинает отмыкание со срединного механизма F, идет к верхнему H и кончает отмыкание верхним H. Реже отмыкание начинается снизу и перекидывается к средине и кверху. Еще реже отмыкание начинается сверху, перекидывается книзу и кончается срединой.
При сравнении кривых продолжительности работы с двумя
[38]
и тремя механизмами
[39]
обнаруживается, что присоединение лишнего (самозамыкающегося) легко отмыкаемого механизма H, расположенного в той же плоскости, что и 2 другие (F и G), в общем не увеличивает среднего срока работы (ввиду предшествующего упражнения этот срок даже меньше прежнего — в особенности при сравнении первых опытов), но способность механизма к самозамыканию, вызывающая необходимость упорядоченного отмыкания, необычайно увеличивает количество излишних движений в единичных опытах, снижает процент идеальных опытов до ничтожной цифры (8%), увеличивает время завершения работы (см. Кривая 1.13 и Кривая 1.14).
При замыкании 3 горизонтальных, находящихся в одной плоскости, разнотипных задвижек R1, O, I (Фото 1.7, Фото 1.8, фиг. 2; Рис. 1.24) также не наблюдается совершенства отмыкания.
В первом опыте изобилуют излишние движения: обезьяна долго не может учесть все три пункта задержки, и только после совершения 8 ненужных движений, после повторного оперирования с верхней и нижней задвижками она обращается к отмыканию средней задвижки (O), кончая работу в 271 сек. времени. Зато в следующих двух опытах обезьяна неизменно начинает с отмыкания именно этой средней задвижки, после чего без промедления отмыкает верхнюю и нижнюю. Время работы и количество излишних движений резко и быстро сокращается, и уже в 3-м опыте обезьяна кончает работу в 18 сек. Но в общем среднее время завершения работы, ввиду малочисленности опытов, довольно велико (123 сек.); оно несколько превосходит сумму среднего времени (112,1 сек.) завершения работы с теми же 3 единично представленными механизмами [40]).
При предъявлении обезьяне 4 задвижек: 3 прежних (R1, O, I) горизонтальных и одной новой — P1 прикрепленной в середине нижнего края дверцы в вертикальном положении, обезьяна совершенно теряется перед новой задачей (Рис. 1.24).

Ее работа хаотична, беспорядочна. Приводимая здесь выписка из протокола
[41]
довольно ярко иллюстрирует порядок оперирования с механизмами в 1-м опыте:
O, Rо, Rз, Rо, Rз, Rо, Rз, I, R, Oо, Oз, Oо, Oз, I, P, R, Iо, R, I, R, I, R, I, Oз, R, отвлечение, Oо, контроль двери, отвлечение, O, R, отвлечение, O, I, P, отвлечение, P, O, R, P, отвлечение, O, R, R, контроль двери, I, R, контроль, R, R, R, P, P, I, O, R, P, I, Iз, Rо, I, Iо, Oо, отвлечение (15 мин. проходит — я указываю на P) — P отмыкает, R.
Из этой выписки явствует, что обезьяна производит 57 обращений к механизмам, вместо 4 необходимых, и все же не справляется с задачей их отмыкания:
Ясно, что наичаще она оперирует с 1-м механизмом, который она многократно повторно отмыкает и замыкает; значительно реже и почти одинаково часто обращается она ко 2-й и 3-й задвижке и всего меньше обращает внимание на эксцентрично расположенную нижнюю — 4-ю — задвижку.
Между делом обезьяна контролирует дверь, отвлекается, снова обращается к оперированию с запорами и снова (вследствие того, что не придерживается никакого порядка при отмыкании) не находит точно места задержки — четвертого механизма (Р), а при кратковременном обращении к нему не улавливает направления его отмыкания. Обезьяна не учитывает и смысла отмыкания и замыкания — она опять производит ряд беспорядочных бесцельных двигательных реакций то здесь, то там, работает наудачу и часто невпопад, лишь контролем двери проверяя возможность освобождения, обращаясь по инерции наичаще к отмыканию 1-й верхней задвижки R и не прилагая достаточного усилия при оперировании с главным препятствием — задвижкой P; вместо этого обезьяна бессмысленно повторно производит замыкание и отмыкание других близрасположенных механизмов.
22 раза она касается 1-й задвижки R в 38,6% случаев 11 раз она касается 2-й задвижки O в 24,5% случаев 13 раз она касается 3-й задвижки I в 22,8% случаев 8 раз она касается 4-й задвижки P в 14,0% случаев
При замыкании только 3 задвижек O, I (горизонтальных, расположенных в средине дверцы, отмыкающихся движением слева направо) и P1 (вертикальной, расположенной по нижнему краю дверцы, отмыкающейся движением сверху вниз, — см. Рис. 1.24 и Фото 1.7, фиг. 2) обнаруживается следующий ход работы (см. табл.).
Во-первых, аналогично предыдущему, в первом опыте обезьяна опять-таки не считает нижнюю задвижку (P) причиной задержки — касается ее попутно, поверхностно и длительно не делает серьезных попыток ее отмыкания.
Только после 4 минут (240 сек.) работы и моего указания на нижнюю задвижку обезьяна производит отмыкание и этой последней.
В последующих 3 — 4 опытах замечается, что обезьяна одной из первых отмыкает именно эту, эксцентрично расположенную вертикальную задвижку, но позднее (начиная с 8-го опыта и по 12-й) обезьяна снова забывает об ее наличности, совершает иногда 20 обращений к двум другим механизмам, прежде чем начать работать с этим последним; она оперирует с последним кратковременно, неудачно, не квалифицирует его, как главный центр задержки, не пытается его отмыкать, а перекидывает свои обследования на 1-й (O) и 2-й (I) механизмы, многократно бесцельно то замыкает, то снова отмыкает эти последние, часто (иногда до 10—11 раз) она контролирует дверь, отвлекается. Обезьяна не обращается к более настойчивому отмыканию этой последней задвижки (P) и после моих многократных указаний на этот механизм. Иногда она начинает работать с другими запорами, вопреки моему указанию на механизм P; если же касается этого последнего, то тянет за центр приложения силы или слишком кратковременно, или применяя несоответствующий способ отмыкания: кусая зубами, ковыряя пальцами, притягивая руками центр приложения силы к себе.
В некоторых случаях (опыт 10) только после 15 минут бесплодной работы, после произведения 25 лишних движений, после моего наглядного отмыкания 3-й задвижки в присутствии обезьяны — животное начинает манипулировать и с механизмом P правильно, словно усваивает нужный прием отмыкания, — но даже и тогда остаются бесцельные движения повторного замыкания механизма после его отмыкания.
В 13-м опыте впервые обезьяна не делает ни одного лишнего обращения к механизмам, по очереди отмыкая 1-й, 2-й, 3-й запоры, в 15 сек. справляясь с их отмыканием; минимальное количество излишних движений в первой серии из 13 опытов равно 3 (в 13-м опыте), максимальное равно 31 (в 8-м опыте; см. табл. и Кривая 1.14).
Общая картина работы обезьяны в первых 13 опытах представляется в следующем виде.
Обезьяна зачастую то забывает об отмыкании эксцентрично расположенного 3-го вертикального запора, то, обращаясь к нему, работает недостаточно энергично, то начинает работать и более настойчиво, но призабывает правильный прием отмыкания.
Все эти причины приводят к тому, что обезьяна производит громадное количество излишних обращений к механизмам (в общем в 5,2 раза больше требуемого
[42],
как и лишних движений контроля двери, чрезмерно задерживается с окончанием работы, тратя в среднем на каждый опыт отмыкания более 4 минут (252,3 сек.), в особо неудачных случаях справляясь с задачей в 15 мин., вопреки тому, что она могла бы при удаче
[43]
и усердии открыть все задвижки в 15 сек.
Следует отметить, что излишние обращения падают главным образом (в 51,5% случаев) на верхнюю (O), наиболее центральную задвижку, наиболее выдающуюся
[44],
в меньшей степени — на среднюю I (26,6%) и в еще меньшей степени — на нижнюю P (21,8). Задвижка № 1 не только наичаще привлекает внимание обезьяны по сравнению с другими двумя механизмами, но она отмыкается в первую очередь. При анализе порядка отмыкания задвижек (см. табл.) обнаруживается, что в подавляющем большинстве случаев 1-е место занимает 1-я задвижка (O) и значительно реже на этом месте встречаются 2-я и 3-я задвижки. Более того, устанавливается даже, что эта 1-я задвижка в преобладающем большинстве случаев (свыше 50%) занимает три первые места и ни разу не отмыкается последней, в то время как задвижка нижняя (P) в 84% случаев отмыкается последней; задвижка средняя (I) по положению занимает все места почти одинаково часто.
В последующих 20 опытах (от 14-го по 33-й) наблюдается значительное усовершенствование обезьяны в деле отмыкания.
Во-первых, ни разу не приходится более напоминать обезьяне о наличности эксцентрично расположенной задвижки № 3 (P), ни разу не обнаруживается забывания приема отмыкания этой последней задвижки.
В связи с этим обстоятельством резко уменьшается (на 100) как количество излишних касаний (которое теперь лишь в 3 раза больше требуемого
[45],
так и среднее время завершения работы (равное 27,4 сек.), укорачивающееся против прежнего более чем в 9 раз
[46].
Соответственным образом уменьшаются в частных единичных случаях цифры максимума и минимума излишних касаний и цифры, выражающие время окончания работы.
Количество излишних движений не превышает 9. В 15% случаев (вместо прежних 7,6%) обезьяна совершенно не делает излишних операций с запорами; несколько уменьшается и количество лишних движений контроля двери.
Что касается вопроса о распределении частоты излишнего оперирования между тремя механизмами (во 2-й серии опытов), то обнаруживается, что обезьяна наичаще начинает обращаться к 3-му механизму (в 46,9% случаев), реже обращение к 1-й задвижке (в 39,3% случаев) и еще реже — ко 2-й (в 13,6% случаев).
При этих повторных манипуляциях обезьяна то отмыкает, то замыкает задвижки, многократно производит бесцельную, излишнюю, никчемную работу.
Обращаясь к анализу порядка отмыкания задвижек, следует отметить, что 3-я задвижка (P) в первую очередь начинает обращать на себя внимание обезьяны (в 45% случаев), реже (в 35% случаев), чем раньше, отмыкается задвижка O и, наконец, еще реже обезьяна начинает свою работу (в 20% случаев) с открывания 2-й задвижки (I).
Задвижка O наичаще (в 60% случаев) отмыкается второй, задвижка P наичаще отмыкается третьей и последней (ею обезьяна и начинает и заканчивает работу), в то время как средняя по положению задвижка (I) почти одинаково часто (в 20 и до 35% случаев) занимает все три места.
Еще значительнее успехи обезьяны сказываются при анализе последних 20 опытов (от 34 по 53).
Количество излишних прикосновений уменьшается на 81 против предыдущего
[47],
на 7 сокращается число движений контролирования двери (равно 10), на 11,1 сек. сокращается среднее время завершения работы (оно равно 16,3 сек. или несколько более 1/4 мин.).
Максимальное количество излишних касаний — 9; цифра аналогична предыдущей, но зато в 30% случаев (вместо прежних 15%) обезьяна совершенно не производит излишних операций. Как и ранее, имеющиеся излишние обращения падают главным образом на 3-й механизм P (в 51,1% случаев), реже и равно часто (в 24,4% случаев) — на механизмы № 1 и 2 (O и I).
Опять меняется номер задвижки, привлекающий на себя первое (как и последнее) внимание обезьяны, — на этот раз таковой является дотоле пренебрегаемая задвижка № 2 (I) (в 45% случаев), 1-я задвижка (O) равно часто пребывает на всех трех местах (и даже на последнем месте), в то время как механизм № 3 (P) отодвигается по порядку его отмыкания на средние места (2-е, 3-е). Вышеприведенная таблица (табл.) показывает наглядно детали этих соотношений.
При сравнении кривой продолжительности работы обезьяны с отмыканием 3 задвижек, расположенных в разных плоскостях (O, I, P) и 3 задвижек в одной плоскости (F, G, H), явствует, что первая кривая, хотя и имеет громадную высоту в своей первой трети, все же она в каждой трети все снижается и снижается, в то время как при наличности самозамыкающейся задвижки кривая обнаруживает в последней трети бо̀льшие подъемы, нежели вначале (сравн. Кривая 1.14 и Кривая 1.14).
Правда, средние сроки завершения работы в первом случае при работе с комбинацией OIP значительно выше, чем в последнем (при работе с комбинацией GFH), но средний процент излишних движений и количество максимума движений у комбинации GFH значительно выше, а более поздние по порядку опыты (12—18) весьма продолжительны. Сравнение работы обезьяны на протяжении всех трех серий опытов по отмыканию 3 задвижек OIP позволяет установить усовершенствование в отношении некоторых сторон ее работы: 1) в отношении усмотрения эксцентрично расположенного механизма P1; 2) в отношении приема отмыкания нового механизма P1; 3) уменьшения количества бесцельных операций; 4) уменьшения срока окончания работы; 5) усвоения некоторого порядка при отмыкании.
Остающиеся дефекты работы: 1) наличность в 70% опытов излишних операций; 2) длительность среднего срока завершения работы вопреки возможности окончания ее в 4 раза более короткий срок; 3) наличность бессмысленных движений замыкания; 4) малое количество идеальных опытов (30%); 5) перемена порядка отмыкания механизмов в каждой новой серии опытов.


Предъявляются механизмы близкой, но несколько измененной, по сравнению с предыдущими, конструкции — задвижки с западанием, требующие произведения двух разных движений в двух разных плоскостях: движения подъема центра приложения силы в вертикальной плоскости снизу вверх и движения отодвигания вбок в горизонтальной плоскости (см. Фото 1.9).
Центры приложения силы в задвижках обладают то большей, то меньшей податливостью и заметностью. Внешний вид задвижек различен.
Задвижка V — черная горизонтальная (Фото 1.9, фиг. 1,2; Рис. 1.25), с легко податливым центром приложения силы, требующим приподнимания вверх и отодвигания вправо.

Обезьяна сразу усматривает центр приложения силы и постигает прием оперирования; она легко и непосредственно производит оба требуемые движения: взявшись одной рукой, смещает центр приложения силы из вертикальной лунки в горизонтальную и движет по горизонтальной лунке то руками, то зубами, тратя на работу 15 сек., быстро сокращая время работы во всех последующих опытах; эпизодические увеличения высоты кривой объяснимы осложнением приема отмыкания: подняв вверх центр приложения силы, обезьяна иногда в несколько приемов движет его в горизонтальной плоскости, оттягивая то руками, то зубами, перехватывая то одной, то другой рукой; иногда она не сразу попадает в горизонтальную лунку, что и удлиняет срок отмыкания.
Первое удачное движение сразу запоминается прочно и производится с каждым новым опытом все более совершенно; усовершенствование выражается в опущении нескольких приемов отодвигания за счет одного, производимого без отрыва, сразу одной и той же рукой и в вертикальной, и в горизонтальной плоскости (что имеет место уже в 5-м опыте, где срок работы укорачивается до 6 сек.); это и отражается на резком спуске кривой времени, особенно к концу сеанса. В первый день работы длительность единичных опытов отмыкания варьирует в пределах 2—22 сек. Средняя длительность опыта 7,8 сек. Последний — 24-й — опыт отмыкания осуществляется в 4 сек.
Усовершенствование в отмыкании этого механизма особенно заметно после 1/2-суточного перерыва во второй день работы с ним: длительность среднего времени завершения работы сокращается в 4,8 раза против таковой 1-го сеанса; пределы вариации длительности единичных опытов — от 1 до 2 сек. Максимальный срок (1-й опыт) — 2 сек.; минимальный — 1 сек.; последний опыт (29) длится 1 сек.; общ. средн. время 6,7 сек. (см. Кривая 1.15).
Механизм W (Фото 1.9, фиг. 1) аналогичной конструкции, прикрепленный в том же горизонтальном положении, видоизмененный по цвету и величине (золотистая задвижка на 2 см меньше предыдущей), отмыкающийся движением спуска центра приложения силы вниз и отодвигания его вправо, преодолевается обезьяной сразу в срок 6 сек.
Как и ранее, обезьяна ухватывается за выступающую пуговицу центра приложения силы то зубами, то руками, спускает ее вниз и движет рукой по горизонтальной лунке вправо. Конечный прием отмыкания аналогичен начальному. Вариация срока окончания работы в пределах от 3 до 8 сек. Последний опыт длится 3 сек. (см. Кривая 1.15).
Среднее время завершения работы с этой последней задвижкой равно 4,4 сек.
Задвижка X1. Прикрепляется механизм той же конструкции и в том же положении — лишь с более крупным сильно выступающим, хотя и с мало податливым пружинящим центром приложения силы, требующим активного удержания при его опускании вниз (в вертикальной плоскости); в горизонтальной плоскости требует движения отодвигания вправо.
Обезьяна сразу применяет нужный прием для отмыкания, прием совершенно аналогичный таковому предшествующих опытов; она не прилагает только в первых своих операциях достаточного усилия при выведении центра приложения силы из вертикальной лунки (сверху вниз) в горизонтальную; центр приложения силы ввиду пружинящих свойств механизма вырывается у нее из рук, поднимается вверх, и это требует от обезьяны повторного многократного (до 3 раз) перехватывания его то руками, то зубами; спускание вниз производится попеременным схватыванием посредством обеих рук, что и увеличивает длительность ее работы в первых 4—5 опытах.
В последующем — с 15-го опыта — обезьяна движет левой рукой в вертикальной плоскости (сверху вниз), правой — в горизонтальной (вправо), но только в 35-м опыте обезьяна усваивает определенную тактику при отмыкании этого механизма: она крепко держит рукой механизм за выступающий центр приложения силы, не выпускает его из рук во время своего действия, вопреки оказываемому им сопротивлению, и отрывается только тогда, когда окончательно отомкнет механизм. Она тянет за центр приложения силы так энергично, что двумя быстрыми движениями одной и той же руки вниз и вбок (чаще правой руки) уже достигает отмыкания, и тогда длительность завершения работы минимальна (от 2 до 5 сек.). Этот прием, как уже было отмечено, устанавливается в самом конце сеанса, а до того нередко обезьяна действует менее активно, берется за центр приложения силы левой рукой, тащит его вниз, перехватывает правой рукой, ведет сначала вниз и в правую сторону; тогда центр приложения силы вырывается у нее, поднимается кверху, и она должна снова и снова (иногда до 5 раз) производить попытки его опускания; движение по горизонтальной лунке не представляет для нее затруднения. Пределы вариации срока работы — от 2 до 42 сек. 1-й опыт отмыкания и самый длительный, последний (30-й) опыт отмыкания продолжается 3 сек. Минимально короткий срок (2 сек.) наступает только в 28-м опыте и повторяется в 29-м.
Средняя длительность работы с этим механизмом в 1-й день оперирования с ним равна 9,9 сек.
Длительность отмыкания значительно сокращается при оперировании с тем же механизмом в следующем сеансе после 3-дневного перерыва в работе; 1-й опыт отмыкания равен всего 6 сек., пределы колебания времени работы — 2—6 сек. Средняя длительность опыта — 3,6 сек., общ. средн. длит. 4,4 сек. (см. Кривая 1.15).

Совсем другие результаты работы обезьяны получаются при предъявлении того же самого механизма в другом положении, именно: не в горизонтальном, а в вертикальном — X2 (Фото 1.7, фиг. 1, 2; Рис. 1.26). В данном случае для отмыкания требуются также два движения отодвигания: вправо (вбок) и вверх, но эти движения идут в иной последовательности и требуют при отмыкании прямо противоположного приема, чем ранее: оттягивание в горизонтальной плоскости вправо — короткое и затрудненное (в силу пружинящего центра приложения силы), отодвигание в вертикальной плоскости вверх — длинное и легкое.
Обезьяна долго не находит центра приложения силы и производит сначала обследования близ колонки, включающей замыкающий стержень, берется за нее, бросает, касается, опять отрывается, дотрагивается зубами, рукой, опять зубами, тянет пальцами за центр приложения силы, отводит, но недостаточно энергично и слишком кратковременно; затем она бросает работу, опять подходит, зубами кусает задвижку, нажимает пальцем сверху, отрывается, грызет снизу, нагибаясь, грызет в средине задвижки, слегка отводит центр приложения силы, но он, вырываясь, возвращается в прежнее положение, обезьяна встает, отводит его направо, отрывается, вертит, опять ведет направо, но не вверх, куда надо. Таким образом, даже найдя руками податливый пункт, она не сразу прилагает достаточно силы, чтобы отвести пуговицу (центр приложения силы) вбок, поэтому, как и ранее, он вырывается у нее и возвращается в прежнее место (в край горизонтальной лунки); после длительного времени работы обезьяна не желает оперировать долее, совершенно игнорируя механизм.
Задача облегчается — центр приложения силы заводится высоко в вертикальную лунку — обезьяне остается сделать одно движение приподнимания вверх; тем не менее обезьяна, чувствуя податливость центра приложения силы вверх и вниз, более тащит вниз, чем вверх.
Делается второе облегчение: центр приложения силы доводится почти до самого верха — одно короткое движение его вверх дает отмыкание. После длительных проб отпирания обезьяна, наконец, улавливает направление податливости после 96 сек. работы; второе приведение механизма в то же положение вызывает более определенное движение оттягивания вверх, ведущее к значительно более скорому (в 16 сек.) отмыканию, причем техника отведения облегчается тем, что обезьяна начинает привставать в вертикальное положение, помогающее ей тянуть вверх сильнее, настойчивее, длительнее. На первое отмыкание механизма всего тратится 112 сек. времени работы.
Осложнение задачи — полное замыкание механизма — после того как животное приучилось находить центр приложения силы и имело опыт удачного отмыкания уже не представляет затруднения, — обезьяна сразу берется за пуговицу, применяет удачное движение оттягивания вбок, повторно настойчиво возобновляет это движение при возвращении центра приложения силы на старое место (ввиду слабого удерживания обезьяной этого центра приложения силы), выводит центр приложения силы из горизонтальной лунки в вертикальную, встает на ноги и тогда движет его вверх, оканчивая работу в 10 сек.
Последующие отмыкания, короткие по времени, явно свидетельствуют о полном постижении обезьяной техники отмыкания этого механизма. Уже в 9-м опыте обезьяна одним движением оттягивает центр приложения силы вправо, вторым отодвигает вверх.
Все же средняя длительность завершения работы с этим последним механизмом довольно высока (20,1 сек.); она более чем в 2 раза превосходит таковую при оперировании с тем же механизмом, но прибитым в горизонтальном положении. Пределы колебания длительности единичных опытов 2—112 сек. 1-й опыт — и самый длительный, последний — равен 5 сек.; минимально короткий опыт — 7-й по счету (см. Кривая 1.15).
Механизм аналогичной конструкции Y1, прикрепленный, так же как V и W, в горизонтальном положении, иной по виду, но с легкоподатливым центром приложения силы, представляющим собою не пуговицу, а кольцо (с оттянутым кверху шпицем), плотно прилегающее к другому кольцу, неподвижно слитому с колонкой внешнего центрального чехла механизма, постигается обезьяной вполне самостоятельно и скоро (Фото 1.9, фиг. 1, 2; Рис. 1.27).
Быстро, после нескольких осязательных проб руками и зубами близ центра приложения силы, обезьяна находит легко падающее вниз кольцо, нащупывающим движением она отыскивает и направление податливости — вниз и в сторону вправо, отодвигая центр приложения силы по горизонтальной лунке в три приема рук, после чего легким нажимом на дверь получает освобождение.
Последующие манипуляции с тем же механизмом еще более уверенны и кратки — уже в 4-м опыте обезьяна фиксирует прием работы, применяя для отмыкания обе руки: левой она спускает центр приложения силы вниз, правой тянет его вправо. Время работы колеблется всего в пределах 3—15 сек.; 1-й опыт — самый длительный, минимально короткий опыт — 5-й по счету, он же и последний. Среднее время окончания работы — 6,8 сек.
Повторение манипуляций с тем же механизмом после перерыва в работе в 23 дня (1-й перерыв), в 45 дней (2-й перерыв) не изменяет совершенства работы.
После первого перерыва — 1-й опыт отмыкания 6 сек., последний — 2 сек. Пределы вариации длительности единичных опытов 2—6 сек. Минимальный срок (2 сек.) повторяется в 4 последних опытах, среднее время окончания работы после перерыва равно 2,8 сек. (ниже, чем ранее).
После второго перерыва (в 45 дней) 1-й опыт отмыкания равен 8 сек. Пределы вариации 1—8 сек. Минимальный срок (1 сек.) повторяется в двух последних опытах, среднее время окончания работы несколько выше (3,6 сек.), чем в предыдущих сериях опытов до перерыва (см. Кривая 1.15). Общ. средн. время окончания работы — 4,4 сек.

Тот же самый механизм Y1 прикрепляется в прямо противоположном положении, требует движения не вправо, а влево (установка механизма Y4) результаты работы резко отличны от предыдущих.
Обезьяна, легко найдя колечко — центр приложения силы, двигает его по привычке длительно вправо, потом, ввиду тщетности попыток отодвигания, пробует новое направление — влево и тогда получает освобождение после 50 сек. работы.
Но прием отмыкания не уловлен, во 2-м опыте (самом длительном в ряду = 220 сек.) обезьяна прикладывается зубами, скребет задвижку, потом ухватывает руками за центр приложения силы, оттягивая его то вправо, то влево; движением влево, отрываясь, в три приема, она производит отмыкание, работая правой рукой; отомкнув, она забывает толкать дверь и производит вторичное замыкание и вторичное отмыкание, увеличивая срок окончания работы.
Фототаблица 1.6. Операции с крюками, задвижками и щеколдами
Фиг. 1. Отпирание крюка Б из комбинации 8 крюков
Но уже в 3-м по счету опыте срок работы резко сокращается, хотя прием отмыкания еще осложнен излишними движениями: три раза обезьяна то отмыкает, то замыкает задвижку.
Время работы неустойчиво колеблется ввиду того, что обезьяна учитывает отомкнутость по звуку щелканья задвижки, и когда этого щелканья не слышит (так как оно бывает лишь при резком движении), она медлит с толканием двери, чем и удлиняет срок окончания работы.
Кривая 1.15. Задвижки V, W, X1, X2, Y1, Y4
Кривая 31. Задвижка сложная V, горизонтальная.
Только в 12-м опыте фиксируется прием работы, и обезьяна даже при отсутствии щелкающего звука производит немедленное толкание двери.
Пределы колебания срока работы от 1,5 до 220 сек. Минимальный срок работы наступает в 10-м и повторяется в 13-м, 17-м, 18-м и последнем опытах (см. Кривая 1.15).
Среднее время окончания работы 24,2 сек.
Фиг. 2. Контроль двери при оперировании с комбинацией из 8 крюков
Фиг. 3. Отмыкание самозамыкающейся щеколды L (из комбинации KLMN)
Фиг. 4. Отмыкание штифтовой задвижки F

Кривая 32. Задвижка сложная W, горизонтальная.
Кривая 33. Задвижка сложная пружинящая X1, горизонтальная.
Кривая 34. Задвижка сложная пружинящая X2, вертикальная.
Кривая 35. Задвижка Y1, горизонтальная, направо.
Кривая 36. Задвижка Y4, горизонтальная, налево.
При одновременном замыкании всех 4 сложных задвижек (Фото 1.9, фиг. 1, 2; Рис. 1.28), именно —
| верхней, вертикальной, отмыкающейся движением сверху вниз, № 1, V1 |
| средней, горизонтальной, отмыкающейся движением слева направо, № 2, Y1 |
| средней, горизонтальной, отмыкающейся движением справа налево, № 3, W1 |
| нижней, вертикальной пруж., отмыкающейся движением снизу вверх, № 4, X2 |
наблюдается следующая картина работы обезьяны (см. табл.).
Рисунок 1.28. Комбинация из 4 сложных задвижек V1, W1, X2, Y1
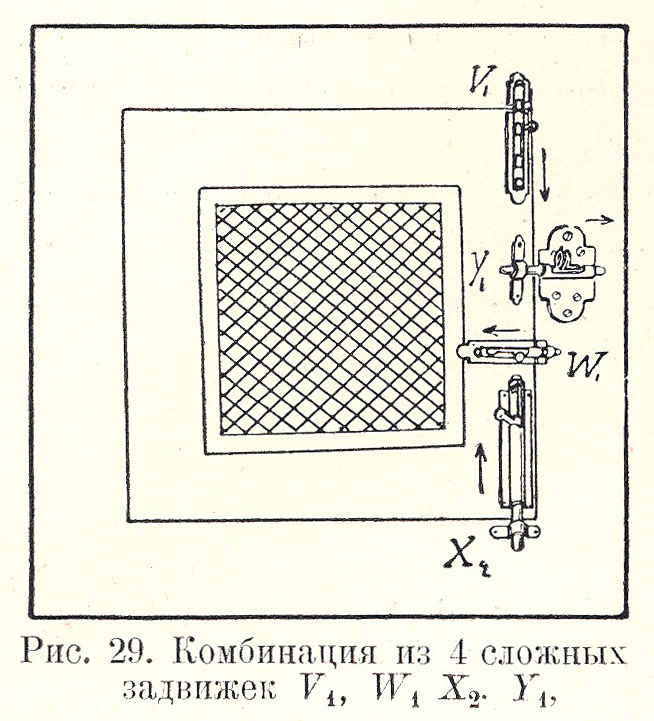
Комбинация из 4 сложных задвижек V1, W1, X2, Y1 (направо, налево, вверх, вниз).
Первоначально, в ближайших трех опытах, обезьяна справляется с разрешением задачи с помощью экспериментатора в длительный срок времени, (от 317 до 540 сек.).
Она забывает о необходимости отмыкать эксцентрично расположенные верхнюю (1) и нижнюю (4) задвижку и многократно бесцельно оперирует с центрально расположенными задвижками (2 и особенно 3). Она обращается к отмыканию удаленных задвижек (1 и 4) лишь после указания на них, но и тогда еще не сразу употребляет достаточно энергичный прием для отмыкания пружинящей (4) задвижки и потому открывает ее после многократного, прерывающегося (15 раз) оперирования.
Уже начиная с 4-го опыта и во всех последующих 20 экспериментах обезьяна совершенно самостоятельно справляется с отмыканием всех 4 запоров и резко уменьшает срок завершения работы, как и количество лишних движений.
В общем, как то явствует из таблицы, процент излишних операций сравнительно невелик, но и процент идеальных опытов также мал (8,3).
Наибольшее количество излишних операций падает главным образом на пружинящую задвижку, более трудную для отмыкания, и на горизонтальную, центральную (а потому наичаще попадающуюся под руку обезьяне), легко податливую задвижку W1, которую обезьяна попутно обследует, то отмыкая, то замыкая. Меньше всего бесцельно трогается более удаленная 2-я горизонтальная задвижка (Y1) и еще меньше верхняя (V2), об отмыкании которой обезьяна зачастую даже забывает. Есть и лишние движения контроля двери.
Что касается порядка отмыкания, то обезьяна придерживается следующей системы.
В общем наичаще она начинает отмыкание с нижней (4-й) задвижки, переходит к следующей над ней — 3-й, еще выше — ко 2-й и кончает отмыкание верхней (1-й) задвижкой.
Реже обезьяна начинает отмыкание с центральной (3-й) задвижки, но после нее идет в направлении книзу (к 4-й), а потом уже кверху (к 2-й и 1-й).
Еще реже отмыкание начинается со 2-й задвижки, перекидывается кверху (к 1-й) и затем уже книзу (к 3-й и 4-й).
Обезьяна ни разу не начинает отмыкания сверху (с 1-й задвижки), очень редко начинает отмыкание и со 2-й, чаще же в первую очередь отмыкает 3-ю и еще чаще — 4-ю.
Кончает отмыкание обезьяна чаще всего 1-й верхней задвижкой.
Средние по порядку отмыкания места занимают средние задвижки, 2-е место наичаще занимает 3-й механизм, 3-е место — 2-й механизм.
При анализе времени завершения работы оказывается, что среднее время окончания отмыкания 4-х задвижек даже в идеальных опытах (где не тратится лишнего времени на бесцельные операции) продолжительнее, чем в единичных, особо удачных опытах; цифра максимума, во много раз превосходит цифру минимума времени окончания работы в единичных опытах (см. только что приведенную таблицу).
И до самого последнего опыта нет систематического спуска кривой продолжительности работы (см. Кривая 1.16); эта кривая обнаруживает резкое падение лишь в самом начале (в 1-й четверти); после того она мало варьирует, пребывает почти на одном и том же уровне, не обнаруживая заметного снижения. При сравнении среднего срока завершения работы (78,4 сек.) обнаруживается, что этот срок значительно выше среднего срока в идеальных опытах.


[22] Пробным мы называем прерванный самой обезьяной безрезультатный опыт; само собой разумеется, что количество времени, затраченного в этих пробных опытах, берется на учет и причитается к следующему за ним удачному опыту отмыкания.
[23] В 8-м пробном опыте.
[24] В 9-м пробном опыте.
[25] Что позволяет ей тянуть после каждого отрыва с большой силой.
[26] Это дает ей возможность иметь задвижку (во все время работы) на уровне рта и увеличивает силу отведения.
[27] На фотографии положение задвижки иное: она открывается влево.
[28] Отражающееся на спуске кривой.
[29] 11-й «пробный» опыт.
[30] Ход работы по отмыканию единичных механизмов — см. предыдущие страницы.
[31] Эта сумма равна 313,3 сек.
[32] Т. е. опытов с отсутствием лишних движений.
[33] И это вопреки тому, что верхняя задвижка O «открытая», скольжение отодвигаемого горизонтального стерженька механизма видно на всем пути его передвижения.
[34] Встречаются главным образом в первой серии из 15 опытов.
[35] Иногда 4-кратного.
[36] Сумма количеств среднего времени завершения работы с Y и I равняется 90,8 сек.
[37] Второй иди даже первой по порядку отмыкания.
[38] F и G.
[39] F, G, H.
[40] Надо, впрочем, принять во внимание, что задвижка R ранее предлагалась в ином положении — требовала движения отодвигания вправо, а не влево, как теперь.
[41] Буквы R, O, I, P означают названия механизмов (R, O, I, P), буква «з» — акт замыкания; буква «о» — акт отмыкания. В целях облегчения набора, номера при буквах опущены, т. е. напечатано R — вместо R1; P — вместо P1. Аналогичный пропуск будет встречаться иногда и позднее — при описании хода работы.
[42] Именно, делает всего 165 лишних касаний к механизмам; совершает 204 движения вместо 39 необходимых.
[43] Что и имеет место в 13-м опыте.
[44]
| Имеет место 85 лишних обращений к 1-й задвижке |
| Имеет место 44 лишних обращений к 2-й задвижке |
| Имеет место 36 лишних обращений к 3-й задвижке |
[45] Именно, 126 вместо 39; следовательно, 66 лишних касаний к механизмам.
[46] Именно, в 9,2 раза; это время меньше на 224,9 сек. против предыдущего (в 1-й серии опытов).
[47] Именно, имеется лишь 45 излишних касаний. (См. пунктирную линию на Кривая 1.14, выражающую количество излишних движений в единичных опытах).